Theobiology
  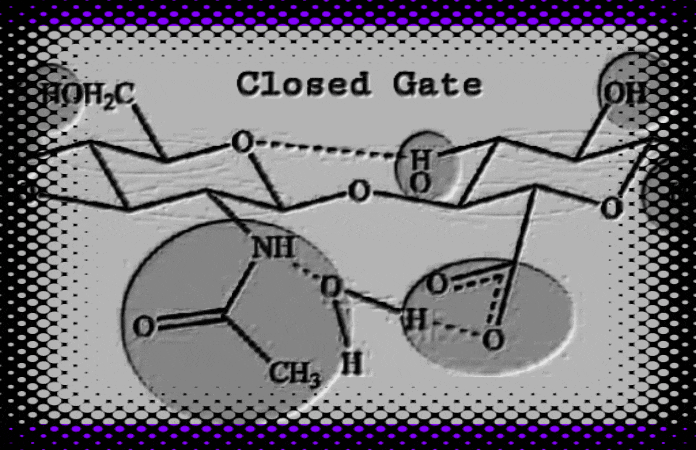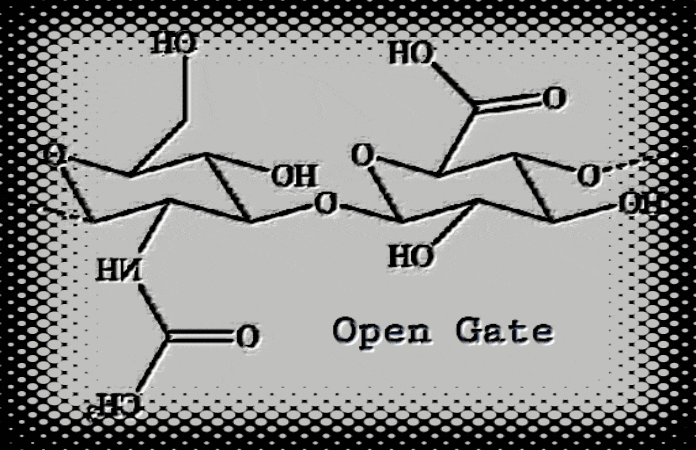
HA, a self-assembling and “gated” double helix shape-shifter, (1, 2, 3, 4) has a biomatrix patterning like that described by Moses 1 (circa 1300 BCE), Ezekiel 2 (circa 600 BCE), and John 3 (circa 96 CE). Hyaluronic acid (HA) biological and physical properties depend on its molecular size (R.E. Turner, P.Y. Lin, M.K. Cowman 1988, and R.E. Turner and M.K. Cowman 1985). In addition, HA is implicated in angiogenesis and chondrogenesis (Toole, B.P.; Turner, R.E., Banerjee, S.D., 1993). During cell matrix engineering, HA facilitates cell migration and promotes cell-cell adhesion through a cell-surface binding protein (R.E. Turner et al, 1991, Toole, B.P., Banerjee, S.D., Turner, R. E., S. Munaim, C. Knudson, 1991). In vitro, hydrated HA fragments can self-assemble into a molecular replica of the Hebrew temple veil and Ezekiel’s wheel (Turner 2021 HaShem Veil, Link). An explanation of theobiology suggests that we can find God in the things that are made (Romans 1:18-23). For instance, if DNA translates God’s instructions, and proteins construct our temple (1Corinthians 3:26), then HA forms the stitches and embroideries within the veil (Psalms 139: 13-14). God created man in his own image and after his likeness (Genesis 1:26). That flesh veil is hyaluronic acid (HA). God writes Strong’s G1449 his letters Strong’s G1989 on the veil of our heart (2 Corinthians 3:3) through his Spirit. Does our biological veil form a quantum interface with the Spirit (Romans 8:16, 1 Corinthians 2:12, and John 4:24)? If the answer is yes, then our hydrated HA veil has quantumness.
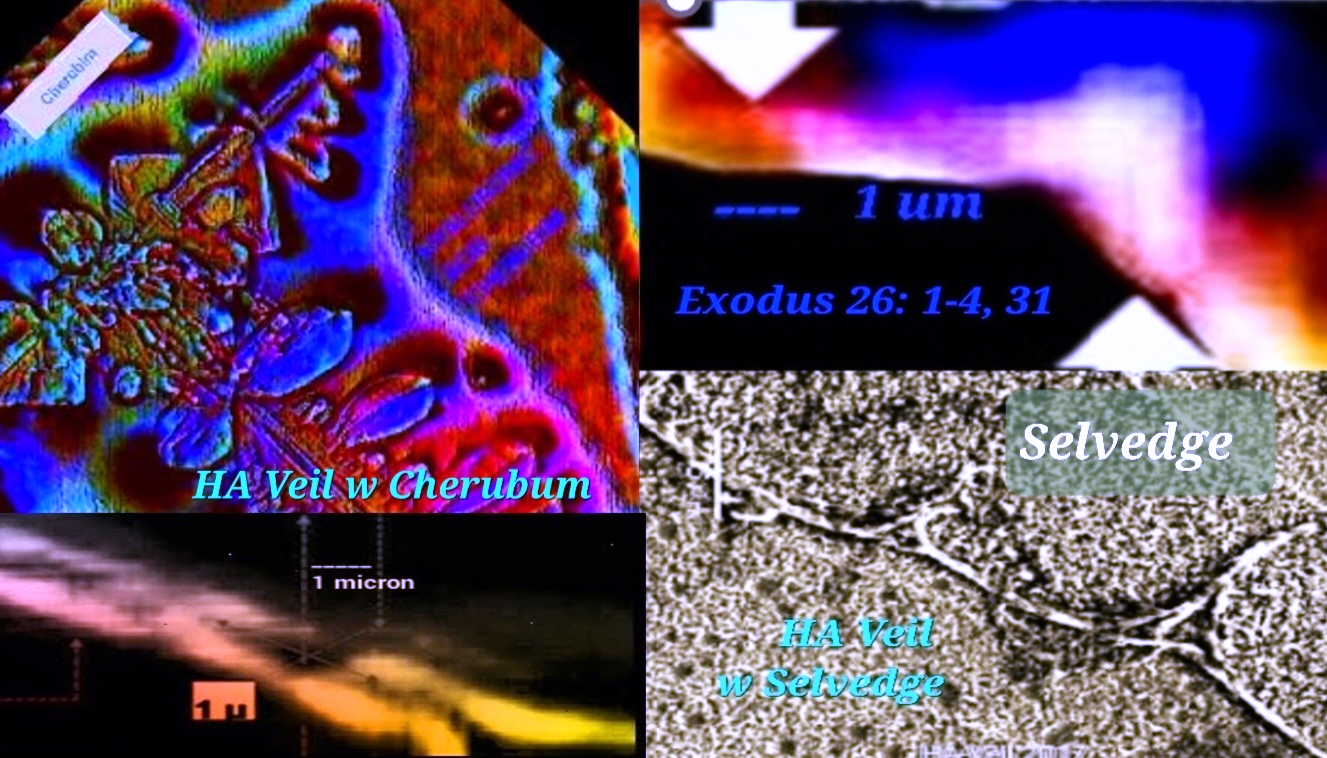
Figure 1. Hyaluronic acid (HA) fragments dehydrating from a water solution on glass slides can self-assemble into nanoscale biomatrix patterning. 4, 5 We see hidden gospel messages such as cherubim and needlework (Exodus 26: 31) embroideries. Evidence for the type of fine twined linen type linkages or selvedges is also found (Exodus 36:11 and Exodus 26:1-4), where the size of the temple veil stitch (28 cubits in length) is given for one curtain. Compare this with the ~28Å in length reported by W.T. Winter et al, 1975. Moses’ human-scale temple veil stitch is compared to HA’s molecular-scale stitch with stunning accuracy. Hydrated HA fragments, nanoBITs (Nanoscale Biocompatible Information Transferons), resemble colorful twisted fibers and exhibit colors such as; blue, purple, red, and white. Ezekiel (Ezekiel 1:4-21) describes “The Veil” (circa 600 BCE) as the “Spirit in the wheels” who came to earth. What Ezekiel saw in a vision matches the HA molecular modeling data reported by a team of non-collaborating researchers.6 Also see the actual HA structures (see figure 6). I made this connection and compared it to my own results on Moses’ description of the veil. Amazingly, these data provide evidence for the Gospel of Jesus Christ. HA is the Spirit-driven biomatrixgenesis veil that covers us and allows spiritual transformation and communication within our biomatrix. That said, the gateway to eternal life is Jesus Christ (Yeshua Ha Mashiach), who is one with the Father (John 10:30). All things consist in him (Colossians 1:17). Created in his image (Genesis 1:27), we are coherent and entangled in him as he is in the Father (John 14:20). Since the veil was torn (Mark 15:38), Believers can enter into his presence through his blood sacrifice.
Figure 2. CLICK IMAGE (GIF). Hyaluronic acid (HA) interconvertible double helix resembles Ezekiel’s wheels (viewed from top) as described by the prophet Ezekiel. Different colors are used to distinguish the characteristics outlined in Ezekiel Chapter 1 relative to Haxaire et al, fig. 6, 2000. Also, HA fragments resembling “fine twined linen, wrought with needlework” (Exodus 26:1-4) revealed hidden gospel messages. The scriptures declare that life of the flesh is in the blood (Leviticus 17:11), but it is the Spirit who gives life (John 6:63). For instance, trees have life, but no blood. The flesh barrier (the veil) connecting the two dimensions is quantum mechanical. “Not until the age of quantum physics did the idea of quantumness in biology emerge.” A quantum gateway most likely connects us, the living temple of God, to the Spirit. On an atomic scale, the HA polyion chains could form an anchor for our soul.7 8 9 10Christ, our righteous veil (Isaiah 61: 10), is the way (1Corinthians 15:54). If we consider the above, we can use AI to generate an image. This image illustrates that we are living souls spiritually entangled while existing within a flesh body.
Figure 3. Turner’s ebook, HaShem Veil, got its title from Hashem, which is the Jewish name for God and also an acronym for Hyaluronic acid Synthases and Hyaluronidases form the Extracellular Matrix. This book highlights the quantumness and the divine patterning of our biomatrix veil, HA. Jesus not only declared himself the temple of God (John 2:18-22), but us as well (1Corinthians 3:16). The flesh molecule, HA, is the molecular patterning of the Hebrew temple veil and Ezekiel’s wheel. In otherwords, man is connected to the spirit dimension through the hierachical self-assembly of his flesh veil, hyaluronic acid (HA). An abundance of hydrogen bonding and water molecules form a polyion-hydrated veil. Within such a veil, our spirit soul becomes entangled. This is why we must be born again from above—of water and of Spirit (John 3:5). Conclusion My laboratory research was conducted at the now Materials Lab (formerly the Center for Materials Science and Engineering) at MIT. I am eternally grateful for that opportunity. My research, which contains theistic overtones, did not provide for collaboration or a peer review process. Post-MIT, my theobiology is focused on hyaluronic acid (HA) as the molecular patterning of the heavenly temple veil. I watched the eclipse of April 8th, 2024 in Fenway, Boston in front of the old Sears’ building. The solar eclipse reminds me that this event may be the last one I will see in my flesh body on earth. The next one, I will watch could be from eternity—on the other side of the veil. |


{kind=link}
{kind=link}